O que significa psychology?
A psicologia é a ciência que estuda o comportamento e os processos mentais humanos. Ela busca compreender como pensamos, sentimos e agimos, além de ajudar na resolução de problemas emocionais e mentais.
O que significa reinforce?
Reforçar significa fortalecer algo que já existe, tornando-o mais resistente e eficaz. Seja na arte do coaching ou na engenharia civil, o ato de reforçar é essencial para garantir a solidez e a durabilidade de qualquer estrutura.
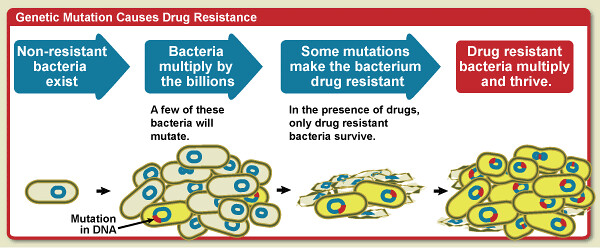
O que significa genetic?
A palavra "genetic" desperta curiosidade e complexidade, evocando um mundo de informações entrelaçadas em nossa essência. Hoje, exploraremos o significado desse termo intrigante e mergulharemos nas raízes da vida e da hereditariedade. Está preparado para descobrir o enigma genético? Vamos aprofundar esse conhecimento!






O que significa transtorno obsessivocompulsivo?
O transtorno obsessivo-compulsivo, conhecido como TOC, é uma condição mental que afeta…










O que significa aproximação na matemática?
A aproximação na matemática é uma poderosa ferramenta que nos permite obter…

O que significa algoritmo na matemática?
Algoritmo na matemática é como uma incrível coreografia numérica, uma dança entre…
O que significa adição?
A adição, um conceito matemático fundamental, revela-se como um enigma estimulante para…

O que significa áreas de figuras geométricas?
Desde os tempos antigos, figuras geométricas têm sido um fascínio para os…
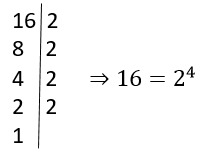
O que significa radiciação?
A radiciação é como um delicado desvendar, um mergulho profundo em busca…


O que significa assunto na matemática?
Matemática, a pedra angular do conhecimento numérico, muitas vezes nos desafia com…

O que significa capitalismo?
O que significa capitalismo? É um conceito que se desdobra em um verdadeiro oceano de interpretações e debates. Desde sua origem na Revolução Industrial até os dias atuais, o capitalismo abrange os pilares da propriedade privada, da livre iniciativa e do mercado competitivo. Mas, será que esse sistema econômico tem se mostrado eficiente e sustentável para todos? Exploraremos essas questões e muito mais neste artigo, mergulhando nas profundezas desse complexo sistema que molda nossa sociedade contemporânea.







O que significa AIDS?
AIDS, uma cama escura onde o medo e a incerteza se encontram. Mas o que realmente significa essa sigla de pesares? Uma resposta complexa, uma doença que assombra a humanidade.…